研究テーマ2:生後脳・成体脳ニューロン新生
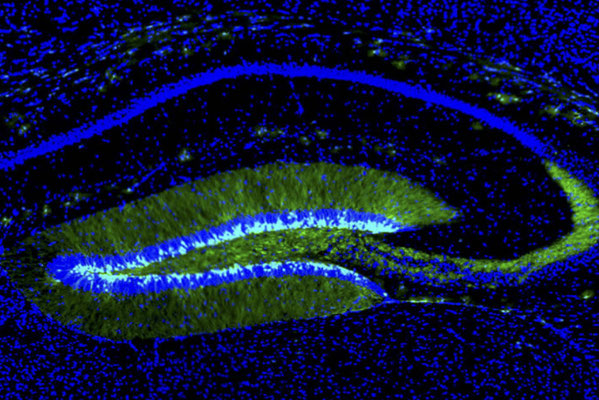
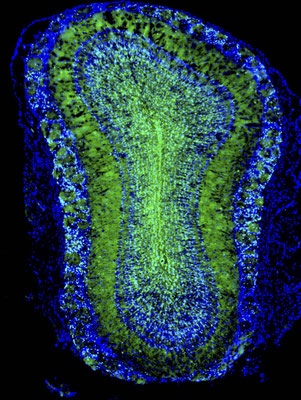
海馬と嗅覚において、蛍光たんぱく質で標識された成体脳で新生したニューロン。
● 成体脳でのニューロン新生の発見
ニューロンの産生は,従来,発生期においてしかおこなわれないと考えられてきました。しかし近年,ヒトを含めた哺乳類の成体の脳においても神経幹細胞が存在し,ニューロンの新生が一生涯続いていることがわかってきました。
1950年代後半,[H3]-チミジンを用いた分裂細胞標識法が開発され,神経解剖学の分野でも使用されるようになりました。Joseph Altman(ジョセフ・アルトマン)らはこの手法を用いて,成体ラット脳のさまざまな部位において,新生ニューロンが存在することを報告しました。1990年後半までには,ヒトも含めて,BrdU取り込み法により調べられたすべての哺乳類においてニューロン新生が観察されました。また,レトロウイルスを用いた細胞系譜追跡実験によってもニューロン新生は確認され,さらに電気生理学的解析により,新生ニューロンは既存の神経回路に組み込まれ,シナプス形成をおこなうことが示されました。
生後脳でのニューロン新生の発見の歴史についてのオリジナル文献の参照は、Joseph AltmanとShirley A Bayer のHPがおすすめです。
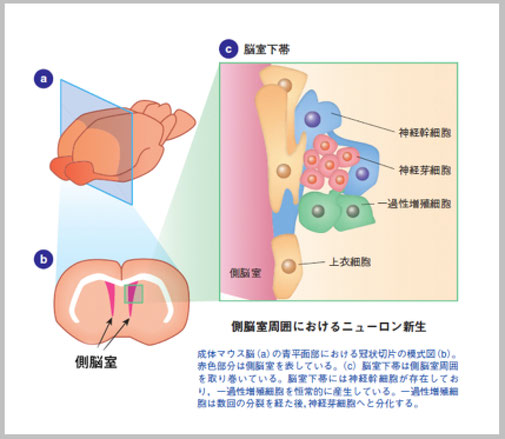
マウスやラットなど齧歯類の成体脳におけるニューロン新生は,主として側脳室の側壁に面した脳室下帯と海馬の歯状回に限局しています。側脳室周囲の脳室下帯で生まれた新生ニューロンは嗅球へと細胞移動し,インターニューロンに分化することがわかっています。脳室下帯は分裂細胞が帯状に連なったもので,側脳室周囲を取り巻いています。脳室下帯には神経幹細胞が存在し,恒常的に新生ニューロンを産生しています。脳室下帯で生まれた新生ニューロンは嗅球へと長い距離を細胞移動し,既存の神経回路に取り込まれます。

成体脳の海馬・歯状回においても,ニューロン新生が恒常的におこっています。顆粒細胞層と歯状回門の間に位置する顆粒細胞下帯とよばれる帯状の領域に神経幹細胞が存在して,顆粒細胞を産生しています。遠く離れた嗅球へと移動する側脳室周囲で産生される新生ニューロンとは異なり,顆粒細胞下帯で新生した顆粒細胞はわずかな距離の細胞移動をおこなうだけで,近接した顆粒細胞層に組み込まれます。
● これからの課題:ニューロン新生を実現するメカニズムとニューロン新生の機能的意義
このようにニューロン新生が一生涯継続するためには、新生ニューロンを生み出す神経幹細胞の増殖・分化・維持が厳密にコントロールされている必要があります。そのような分子メカニズムはまだまだ不明な点が多いです。また、なぜ側脳室周囲の脳室下帯と海馬の歯状回でだけ、神経幹細胞が維持され、ニューロン新生が継続できるか?という点も大きな謎です。
新生ニューロンが組み込まれる嗅球は嗅覚情報を脳内部に伝えるための一次情報処理センターです。また、海馬は脳の記憶中枢と考えられており,その中の歯状回は海馬へ信号が入ってくる際の入り口です。これらの領域でニューロン新生が活発に起きているということは、ニューロン新生が嗅覚機能や、記憶や学習などの高次脳機能に何らかの役割を果たしていると予想されます。実際、ニューロン新生を阻害したマウスを用いて記憶テストを行ったところ、迷路を用いた空間学習テストにおいて,空間記憶は形成されるものの,記憶の長期保持能が野生型マウスよりも著しく低いことがわかりました。このようにニューロン新生は記憶・学習などの高次脳機能に関与していることが分かってきましたが、ニューロン新生は、記憶の同時性・関連性や、不必要な記憶の忘却、情動制御やより本能的な行動の制御など、もっと多様な脳機能に関与している可能性があります。
私たちは、これらの未解決の疑問に対してマウスをモデル生物として用いて解析を行っています。特に、成体脳の神経幹細胞や新生ニューロンを特異的に遺伝子操作できるようなトランスジェニックマウスやノックインマウスを開発して解析を行っています。これらの遺伝子改変マウスを用いて、組織学・生理学や行動学を組み合わせ、成体脳ニューロン新生の全体像と機能的意義の解明、さらにはニューロン新生を用いた脳機能改善への応用に貢献できればと思っています。
代表的な論文:
Li, W.L., Chu, M.W., Wu, A., Suzuki, Y., *Imayoshi, I. and *Komiyama T. (2018) Adult-born neurons facilitate olfactory bulb pattern separation during task engagement. eLife, 7, e33006..
Sakamoto, M., Ieki, N., Miyoshi, G., Mochimaru, D., Miyachi, H., Imura, T., Yamaguchi, M., Fishell, G., Mori, K., Kageyama, R. and Imayoshi, I.* (2014) Continuous postnatal neurogenesis contributes to formation of the olfactory bulb neural circuits and flexible olfactory associative learning. The Journal of Neuroscience 34: 5788-5799.
Sakamoto, M., Imayoshi, I., Ohtsuka, T., Yamaguchi, M., Mori, K. and Kageyama, R. (2011)
Continuous neurogenesis in the adult forebrain is required for innate olfactory responses. PNAS 108: 8479-8484
Imayoshi, I., Sakamoto, M., Yamaguchi, M., Mori, K. and Kageyama, R. (2010) Essential roles of Notch signaling in maintenance of neural stem cells in the developing and adult brains. The Journal of Neuroscience 30: 3489-3498.
Imayoshi, I., Sakamoto, M., Ohtsuka, T., Takao, K., Miyakawa, T., Yamaguchi, M., Mori, K., Ikeda, T., Itohara, S. and Kageyama, R. (2008) Roles of continuous neurogenesis in the structural and functional integrity of the adult forebrain. Nature Neuroscience 11: 1153-1161.